NKduHFKvDJqTniJOiPRFCQZnduSxRVENzvlAfmfidtsdr
LOsqzzYlbUTotjiEZfpcxx
soxoQaHUV
IrlyiyeuDKzCYXbLikppt
|
單細胞測序技術服務 靶向lncRNA单细胞全转录组测序 單細胞全轉錄組測序 |
|
生物分子凝聚體研究 HyPro靶RNA临近标记技术 |
|
RNA-蛋白相互作用 HyPro - MS CHIRP – MS RNA pull-down MS |
RNA-RNA/DNA相互作用 HyPro-seq/芯片 CHIRP-seq |
蛋白-RNA相互作用 AGO APP seq/芯片 RIP-RNA seq/芯片 |
蛋白-蛋白相互作用 CoIP-MS/AP-MS |
|
NGS测序技术服务 DRIPc-seq |
|
NGS测序技术服务 環狀DNA测序 |
|
基因芯片技術服務 Small RNA修饰芯片 m6A单碱基分辨率芯片 mRNA&lncRNA表观转录组芯片 circRNA表观转录组芯片 |
NGS测序技术服务 表觀轉錄組學測序服務 RNA m6A甲基化测序(MeRIP Seq) |
LC-MS mRNA碱基修饰检测 tRNA碱基修饰检测 |
PCR技术服务 MeRIP-PCR技术服务 m6A绝对定量RT-PCR技术服务 m6A单碱基位点PCR(MazF酶切法)技术服务 |
|
NGS测序技术服务 DNA(羟)甲基化测序(抗体法) DNA甲酰基胞嘧啶(5fc)修饰测序 DNA 5hmC 测序(化学法) 染色質免疫共沉澱測序 |
PCR技术服务 MeDIP-qPCR hMeDIP-qPCR ChIP-qPCR |
|
Ribo-seq Ribo seq |
核糖體-新生肽链复合物(RNC) RNC联合 circRNA芯片 RNC联合 lncRNA芯片 RNC联合mRNA-seq |
tRNAs曾經被認爲是高丰度的、广泛存在的、被动参与的mRNA解碼器及蛋白翻譯元件。通過tRNA上的反密碼子與mRNA的密碼子結合,搬运指定的氨基酸以完成蛋白翻译。攜帶同種氨基酸而反密碼子不同的tRNAs被稱爲“isoacceptors”;攜帶同種氨基酸且反密碼子相同,但序列不同的tRNAs則互爲“isodecoders”。最新的科研進展意外地發現tRNA isoacceptors和isodecoders具有特異性的、非被动参与翻译的主动调控功能,可以显著影响生物学过程和疾病进展。此外,tRNAs在體液中高丰度存在的特點使其成爲臨牀應用的生物標誌物。深入探究tRNAs的潛在功能讓這個經典的分子煥發新生。
|
經典教科書中的認知 |
最新前沿進展 |
|
l 普通的,被动参与蛋白翻译的RNA l 高丰度,广泛存在的 |
l 細胞、组织、疾病和时序特异性表达 l Isoacceptors通過擺動解碼效應調控翻譯 l Isodecoders影響生理進程和疾病發生 l tRNAs和tRNA衍生小片段tRF&tiRNA可作爲高丰度的體液檢測生物標誌物 |
細胞、组织和疾病特异性表达
tRNA isoacceptors和 isodecoders的表达谱与细胞分化和增殖密切相关(Fig.1A),在胚胎发育过程中呈现出高度的组织、细胞类型、时间和空间特异性(Fig.1B),而且在腦、胸腺、血液、脾脏、肝脏、睾丸、卵巢和乳腺癌中有疾病特异性表达(Fig.1C)。這種表達特異性與生物學功能和病理生理學緊密關聯,赋予tRNA更好的生物标志物潜力。
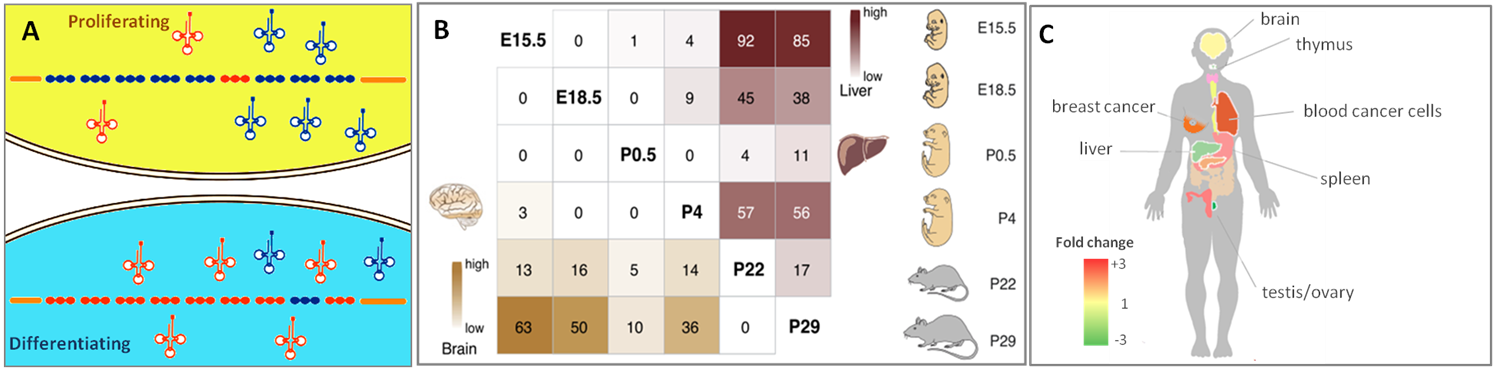
圖1.(A)增殖和分化狀態的細胞有不同的tRNA表達譜特徵[1]。(B)tRNAs具有組織、器官、发育阶段特异性表达[2]。(C) tRNAs在不同人體組織和疾病中差異表達。例如骨髓瘤细胞相比于正常骨髓细胞通常具有更高的核tRNA表達[3]。
Isoacceptors的擺動解碼(wobble decoding)影響蛋白翻譯
攜帶每種氨基酸的tRNA isoacceptor家族組成了tRNA庫,参考该tRNA庫可以更好地研究tRNA的密碼子使用偏好、蛋白翻译效率和精确性、生物学过程及其在人类疾病中的作用等。密码子可以被完全互补配对的同工反密码子解码,或者被在摆动解码位置有单碱基错配的近似同工反密码子解码。tRNA庫中攜帶同工/近似同工反密碼子tRNA的比例可被動態調控從而影響翻譯效率、準確性和轉錄本穩定性(Fig.2)[4]。在各種條件下,tRNA庫會發生改變來維持穩態或促進特定的基因表達進程[5]。Isoacceptor表達水平的紊亂可導致癌症(Fig.3A)[6]和神經退行性疾病(Fig.3B)[7]等疾病的发生。
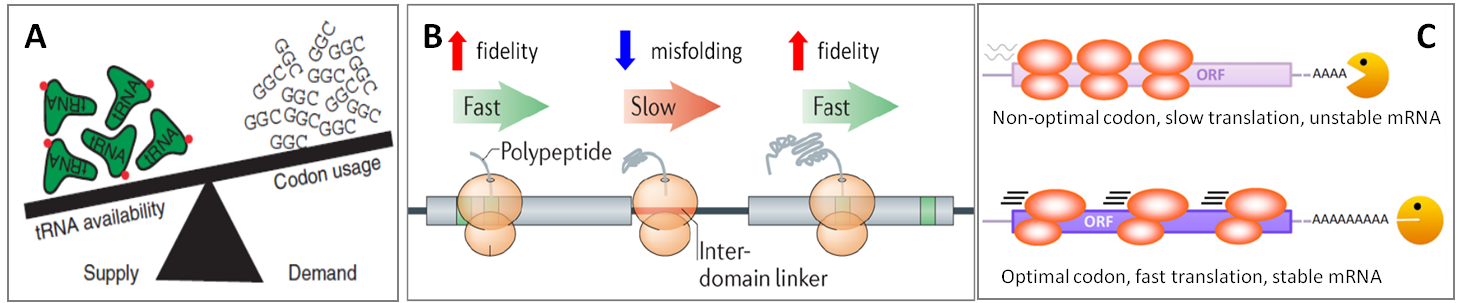
圖2.(A)當同工tRNA供應量超過蛋白翻譯需求量時稱爲最優密碼子。当同工tRNA供應不足或需要使用近似同工tRNA時稱爲非最優密碼子。(B)最優密碼子加快翻譯速率並提高解碼精確性。非最优密码子翻译速率慢得多但有利于蛋白正确折叠,通常发生在编码的蛋白结构域之间的连接区域。(C)經過最優密碼子時更快的翻譯延伸速率可保護mRNA不被降解。使用非最优密码子导致翻译速率降低、核糖体停滞、暴露的mRNA更容易降解,从而缩短mRNA半衰期。
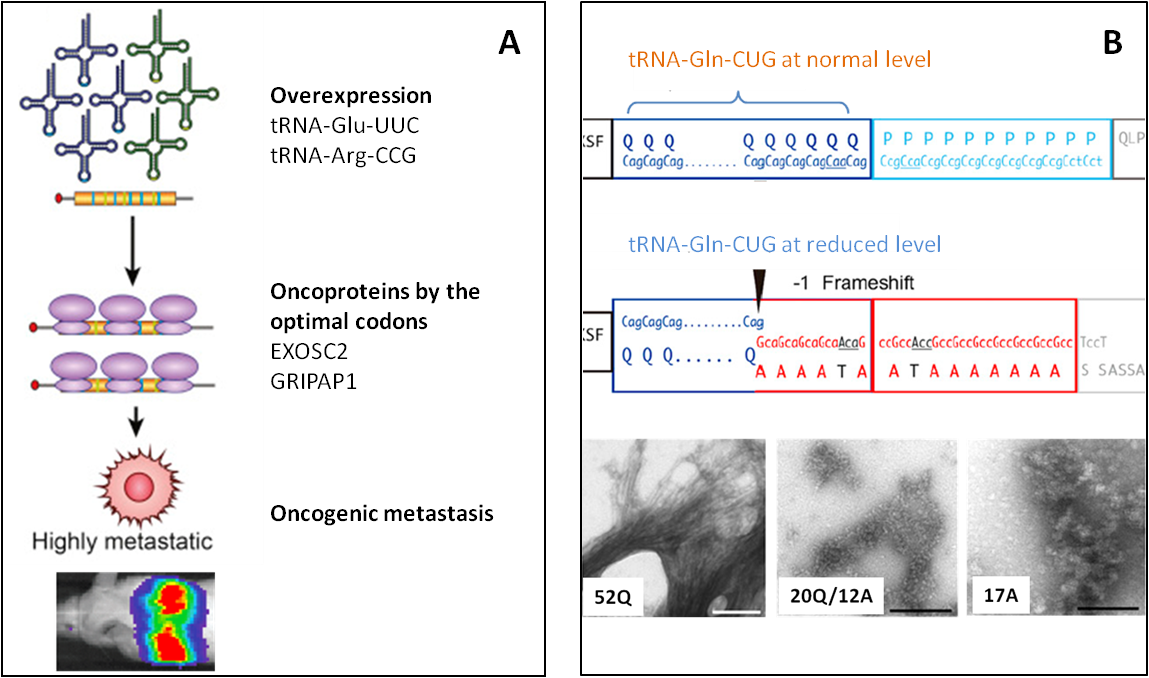
圖3.(A)过表达tRNA-Glu-UUC和tRNA-Arg-CCG可上调以它们为最优密码子的原癌蛋白EXOSC2和GRIPAP1的翻譯,促进原癌基因驱动的乳腺癌侵袭过程[6]。(B) 正常供应量的tRNA-Gln-CUG可以正確翻譯亨廷頓蛋白中的多聚谷氨醯胺鏈PolyQ重複區域,tRNA-Gln-CUG含量降低則會導致移碼翻譯(编码Gln的密碼子CAG閱讀框前移一位變爲編碼Ala的密碼子GCA)产生错误的多聚丙氨酸链PolyA。亨廷顿蛋白中Q/A的比率決定了蛋白聚集狀態(52Q正常狀態,20Q/12A中間態,17A致病聚集狀態)从而影响亨廷顿舞蹈症的严重程度[7]。
Isodecoders影響生物學過程和疾病發生
高度保守的tRNA isodecoder序列可呈現不同的組織特異性表達,其发挥的功能也具有明显差异。例如,小鼠的5種tRNA-Arg-UCU isodecoders只有一種在中樞神經系統(CNS)中特異性表達(Fig.4A)。它的缺失會引發mRNA上核糖體停滯,ATF4驅動的壓力應激反應和廣泛的神經變性(Fig.4B, C, D)。因此CNS特異性表達的isodecoder具有維持神經元細胞穩態和防止神經變性的新功能[8]。
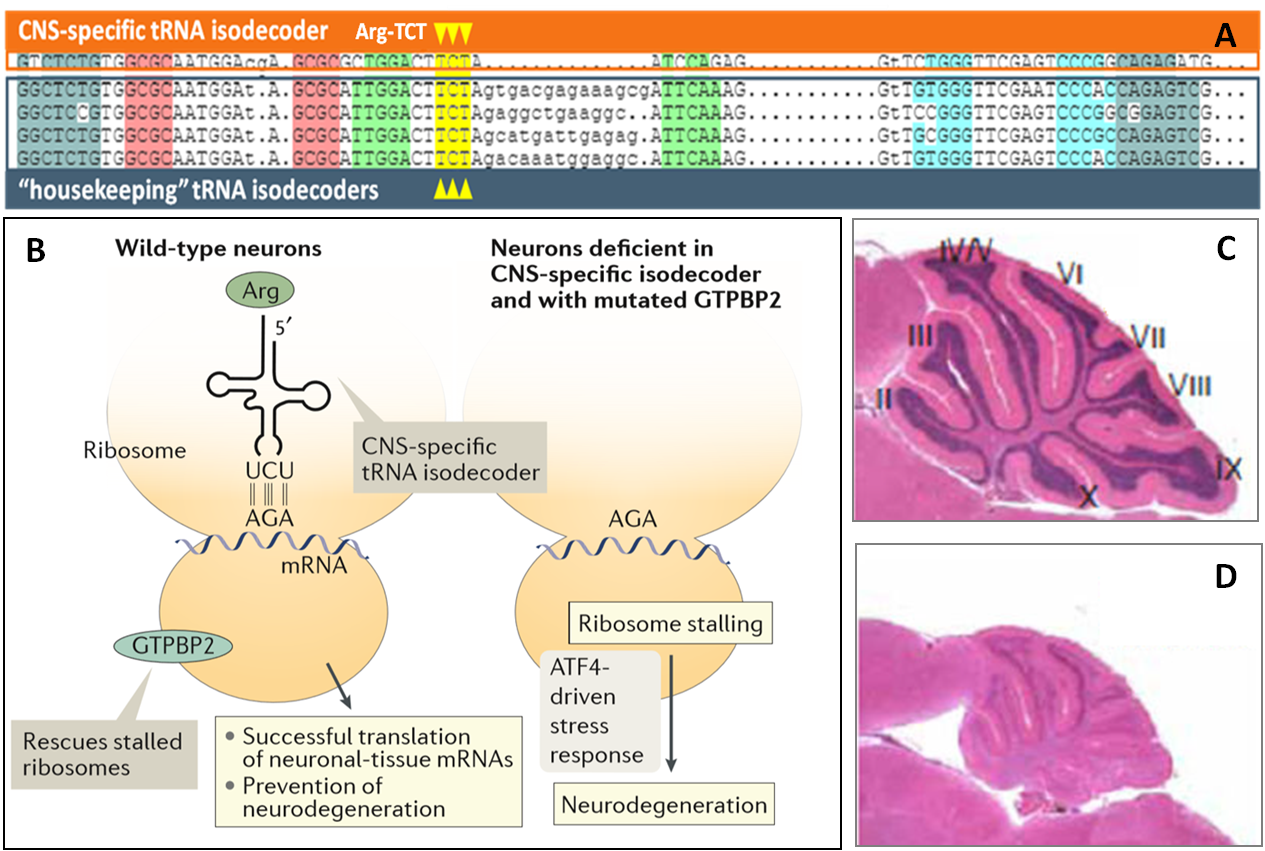
图4. (A) CNS特異性表達以及“管家”tRNA-Arg-UCU isodecoder基因序列 (B) CNS特異性表達的isodecoder对于正常神经元的翻译顺利进行至关重要,否则,该细胞将遭受神经变性。(C) 正常野生型脑。 (D) 缺失CNS特異性tRNA-Arg-UCU isodecoder 的神经变性脑 [8].
非經典、非翻译元件的调控型tRNA功能
tRNA可以通過不直接參與蛋白翻譯的方式調控細胞功能。例如,tRNA可以結合並抑制細胞色素C在凋亡過程中形成凋亡複合體 (Fig. 5)[9,10]。因此,tRNA可作爲凋亡抑制因子。
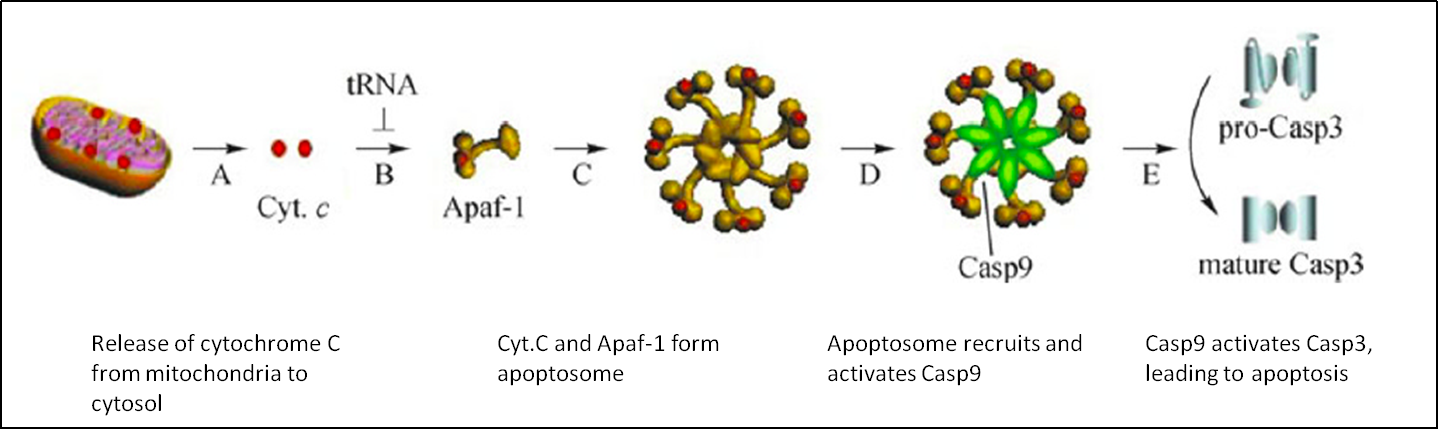
圖5. tRNA結合細胞色素C,抑制凋亡复合体形成,进而抑制细胞凋亡[9,10]。
在熱休克、缺氧或营养剥夺等多种细胞压力下,tRNA也是最早的應答分子,通过多种调控机制关闭整个蛋白翻译(Fig. 6)[11]。
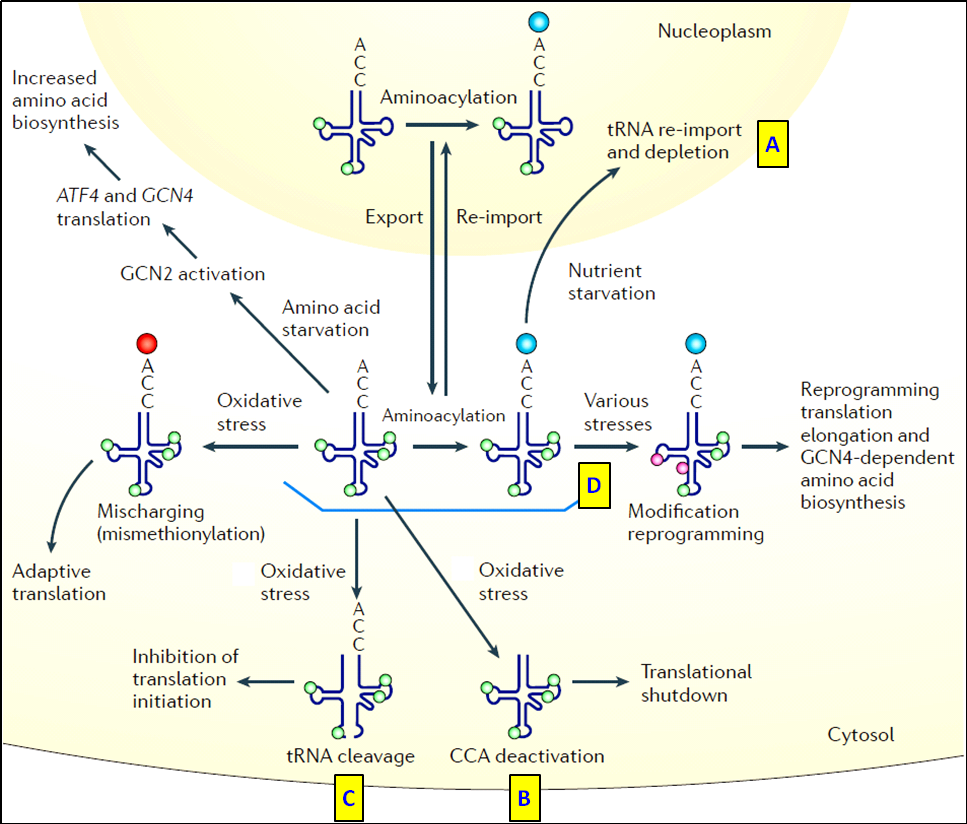
图6.响应细胞压力应激的tRNAs (A) 营养缺乏导致tRNAs從胞質重新運回胞核,使胞质中参与蛋白翻译的tRNA庫耗竭; (B) tRNA 3’CCA 臂失活; (C) 缺氧诱导tRNA斷裂形成tRF和tiRNA片段,抑制蛋白翻译起始。(D)在營養匱乏期間,tRNA修飾重編程促進氨基酸合成途徑中蛋白質/酶的翻譯[11]。
tRNA作爲體液檢測的生物標誌物
小RNA(例如microRNA)已被廣泛地用作生物標誌物。由于tRNA的很多理想特徵,它们现在正成为一类新型的生物标志物。它们是高丰度表达的小RNA,在某些体液样本中含量甚至高于miRNAs(Fig. 7A)[12, 13]。tRNA片段大量富集於神經元細胞外泌體中,成为仅次于rRNA的第二豐富RNA種類,含量远超miRNA (Fig. 7B)[14]。
高丰度、被大量修饰保护的稳定性、体液样本中富集的特点意味着tRNA有更好的生物標誌物敏感度。加上tRNA差異表達與疾病的緊密關聯(Fig. 1),tRNA可以成爲一類具有更大潛力的生物標誌物。
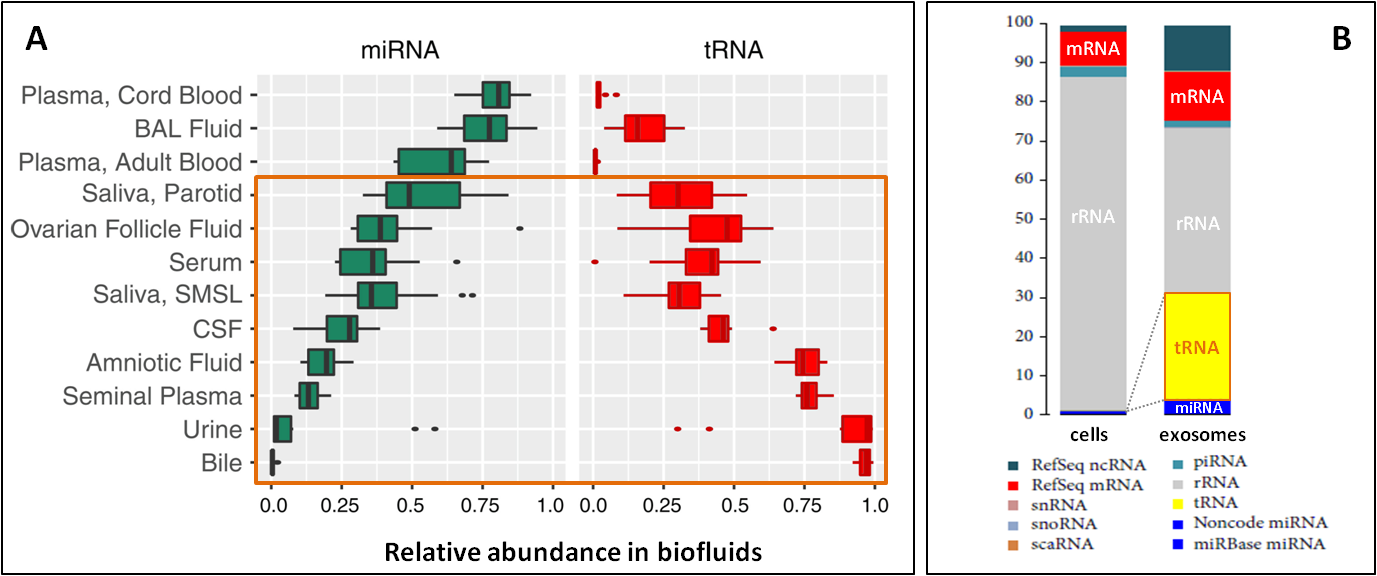
圖7.(A)miRNA與tRNA在體液中相對比例的對比,在某些类型体液中tRNA含量高於miRNA [12, 13]。(B)與其他胞內RNA相比,tRNA在外泌體中高度富集,远超miRNA[14]。
參考文獻
[1] Gingold H. et al. (2014) Cell [PMID: 25215487]
[2] Schmitt B.M. et al. (2014) Genome Res. [PMID: 25122613]
[3] Geslain R. and G. Eriani (2014) Translation [PMID: 26779404]
[4] Hanson G. and J. Coller (2018) Nat. Rev. Mol. Cell Biol. [PMID: 29018283]
[5] Gavin H. et al. (2014) Molecular Cell Biology [PMID: 29018283]
[6] Goodarzi H. et al. (2016) Cell [PMID: 27259150]
[7] Girstmair H. et al. (2013) Cell Rep [PMID: 23352662]
[8] Ishimura R. et al. (2014) Science [PMID: 25061210]
[9] Mei Y. et al. (2010) Mol. Cell [PMID: 20227371]
[10] Mei Y. et al. (2010) Protein Cell [PMID: 21113408]
[11] Kirchner S. and Z. Ignatova (2015) Nat. Rev. Genet. [PMID: 25534324]
[12] Godoy P.M. et al. (2018) Cell Rep [PMID: 30380423]
[13] Schageman J. et al. (2013) Biomed Res Int [PMID: 24205503]
[14] Quek C. et al. (2017) RNA Biol [PMID: 28005467]
给我免费播放片高清在线观看生物丨数谱生物 tRNA研究技術平臺
tRNA 表達
Arraystar nrStar™ tRNA PCR 芯片(H/M)
分別檢測185個人/小鼠 tRNAs,覆盖GtRNAdb數據庫所有的細胞核反密碼子
(1)從tRNA isoacceptors和isodecoders兩個層面進行表達水平分析:
·Isoacceptor水平檢測具有相同反密碼子的總體isodecoder水平,适用于tRNA表達譜分析
·Isodecoder水平檢測單獨tRNA isodecoders,适用于分析isodecoder特異性的功能
芯片包含所有的反密碼子,可获得同源和近似同源tRNA的表達信息,从而分析二者比值在生物和疾病条件下的变化。
(2)tRNA去甲基化處理,去除了阻碍tRNA反轉錄過程的甲基化修飾,顯著提高檢測靈敏性
完善的tRNA前處理方案,去除tRNA上的甲基化修飾和未端氨基對測序的阻礙,精确检测全部tRNA
tRNA形成
Arraystar NuRNATM small RNA Biogenesis Proteins PCR芯片
檢測185個人 小RNA( miRNA、tRNA、 piRNA等)生物合成通路上的酶和蛋白因子
tRNA修飾
Arraystar NuRNATM tRNA Modification Enzymes PCR芯片
檢測85個在tRNA修飾過程中起關鍵作用的酶和蛋白因子
利用液相色譜-質譜聯用技術分析和檢測tRNA上的核酸修飾


